Untuk dapat berbicara secara komprehensif tentang hemoglobin (Hb), ada baiknya berurusan terlebih dahulu dengan mioglobin (Mb) yang sangat mirip dengan hemoglobin tetapi jauh lebih sederhana. Ada hubungan kekerabatan yang ketat antara hemoglobin dan mioglobin: keduanya merupakan protein terkonjugasi dan kelompok prostetiknya (bagian non-protein) adalah kelompok heme .
Myoglobin adalah protein globular yang terdiri dari rantai tunggal sekitar seratus lima puluh asam amino (tergantung pada organisme) dan berat molekulnya sekitar 18 Kd.
Seperti disebutkan, ia dilengkapi dengan kelompok heme yang dimasukkan dalam bagian hidrofobik (atau lipofilik) dari protein, yang terdiri dari lipatan yang disebabkan oleh struktur heliks α dari protein berserat.
Myoglobin terutama terdiri dari segmen heliks α, hadir dalam jumlah delapan dan terdiri, hampir secara eksklusif, residu non-polar (leusin, valin, metionin dan fenilalanin) sementara residu polar praktis tidak ada (asam aspartat, asam glutamat, lisin dan arginin); satu-satunya residu kutub adalah dua histidin, yang memainkan peran mendasar dalam serangan oksigen ke kelompok heme.
Grup heme adalah kelompok kromofor (menyerap dalam kasat mata) dan merupakan kelompok fungsional mioglobin.
Lihat juga: hemoglobin terglikasi - hemoglobin dalam urin
Sedikit chemistry
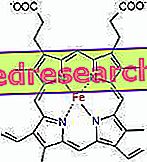
Hubungan antara protoporphyrin dan besi adalah ikatan khas dari apa yang disebut senyawa koordinasi yang merupakan senyawa kimia di mana atom pusat (atau ion) membentuk ikatan dengan spesies kimia lain dalam jumlah yang lebih tinggi dari pada bilangan oksidasi (muatan listrik). Dalam kasus heme, ikatan ini bersifat reversibel dan lemah.
Jumlah koordinasi (jumlah ikatan koordinasi) dari besi adalah enam: mungkin ada enam molekul di sekitar besi yang berbagi elektron ikatan.
Untuk membentuk senyawa koordinasi, diperlukan dua orbital dengan orientasi yang benar: satu dapat "membeli" elektron dan yang lain mampu menyumbangkannya
Dalam heme, zat besi membentuk empat ikatan planar dengan empat atom nitrogen yang berada di tengah cincin proto-porfirin dan ikatan kelima dengan nitrogen histidin proksimal; besi memiliki tautan koordinasi bebas keenam dan dapat mengikat oksigen.
Ketika besi dalam bentuk ion bebas, orbital tipe-dnya memiliki energi yang sama; dalam mioglobin, ion besi terikat pada protoporphyrin dan histidin: spesies ini secara magnetis mengganggu orbital besi; luasnya gangguan akan berbeda untuk berbagai orbital sesuai dengan orientasi spasial dan spesies yang mengganggu. Karena energi total dari orbital harus kembali ke konstan, perturbasi menyebabkan pemisahan energik antara berbagai orbital: energi yang diperoleh oleh beberapa orbital, setara dengan energi yang hilang oleh yang lain.
Jika pemisahan yang terjadi antara orbital tidak terlalu besar, pengaturan elektronik putaran tinggi lebih disukai: elektron ikatan mencoba untuk mengatur diri mereka sendiri dalam putaran paralel dalam beberapa kemungkinan sub-level (multiplisitas maksimum); jika, di sisi lain, gangguan sangat kuat dan ada pemisahan besar antara orbital, mungkin lebih mudah untuk memasangkan elektron ikatan di orbital energi yang lebih rendah (putaran rendah).
Ketika besi berikatan dengan oksigen, molekul mengambil disposisi putaran rendah sedangkan ketika besi memiliki hubungan koordinasi bebas keenam, molekul memiliki putaran tinggi.
Berkat perbedaan putaran ini, melalui analisis spektral mioglobin, kami dapat memahami apakah ada oksigen terikat (MbO2) atau tidak (Mb).
Myoglobin adalah protein khas otot (tetapi tidak hanya ditemukan di otot).
Myoglobin diekstraksi dari paus sperma di mana ia hadir dalam jumlah besar dan kemudian dimurnikan.
Cetacea memiliki respirasi seperti manusia: memiliki paru-paru mereka harus menyerap udara melalui proses pernapasan; paus sperma harus membawa oksigen sebanyak mungkin di otot-otot yang dapat mengakumulasi oksigen dengan mengikatnya ke mioglobin yang ada di dalamnya; Oksigen kemudian dilepaskan perlahan ketika cetacean terbenam karena metabolismenya membutuhkan oksigen: semakin besar jumlah oksigen yang dapat diserap paus paus dan semakin banyak oksigen yang tersedia selama penyelaman.

Untuk membuat daging lebih atau kurang merah adalah isi dari hemoprotein (itu adalah heme yang membuat daging menjadi merah).
Hemoglobin memiliki banyak analogi struktural dengan mioglobin dan mampu mengikat oksigen molekuler secara reversibel; tetapi, sementara mioglobin terbatas pada otot dan jaringan perifer pada umumnya, hemoglobin ditemukan dalam eritrosit atau sel darah merah (mereka adalah sel pseudo, yaitu, mereka bukan sel nyata) yang membentuk 40% dari darah.
Hemoglobin adalah tetrameter, yang terdiri dari empat rantai polipeptida, masing-masing dengan kelompok heme dan dua per dua (pada manusia ada dua rantai alfa dan dua rantai beta).
Fungsi utama hemoglobin adalah transportasi oksigen ; fungsi lain dari darah di mana hemoglobin terlibat adalah pengangkutan zat ke jaringan.
Di jalur dari paru-paru (kaya oksigen) ke jaringan, hemoglobin membawa oksigen (pada saat yang sama zat lain juga mencapai jaringan) sementara di jalur sebaliknya, ia membawa serta limbah yang dikumpulkan dari jaringan, terutama karbon dioksida yang dihasilkan dalam metabolisme.
Dalam perkembangan manusia ada gen yang diekspresikan hanya untuk jangka waktu tertentu; untuk alasan ini kami memiliki hemoglobin yang berbeda: janin, embrionik, dari pria dewasa.
Rantai yang membentuk hemoglobin yang berbeda ini memiliki struktur yang berbeda tetapi dengan kesamaan fungsi yang mereka lakukan kurang lebih sama.
Penjelasan tentang keberadaan beberapa rantai berbeda adalah sebagai berikut: selama proses evolusi organisme, hemoglobin juga telah berevolusi mengkhususkan diri dalam pengangkutan oksigen dari daerah yang kaya akan daerah kekurangan. Pada awal rantai evolusi, hemoglobin membawa oksigen dalam organisme kecil; dalam perjalanan evolusi organisme telah mencapai dimensi yang lebih besar, oleh karena itu hemoglobin telah berubah untuk dapat mengangkut oksigen ke daerah yang lebih jauh dari titik ia kaya; untuk melakukan ini, struktur baru rantai yang merupakan hemoglobin telah dikodifikasikan dalam proses evolusi.
Myoglobin mengikat oksigen bahkan pada tekanan sederhana; dalam jaringan perifer ada tekanan (PO2) sekitar 30 mmHg: pada tekanan ini mioglobin tidak melepaskan oksigen, sehingga tidak akan efektif sebagai pembawa oksigen. Sebaliknya, hemoglobin memiliki perilaku yang lebih elastis: ia mengikat oksigen pada tekanan tinggi dan melepaskannya ketika tekanan berkurang.
Ketika protein aktif secara fungsional, ia dapat sedikit mengubah bentuknya; misalnya, mioglobin teroksigenasi memiliki bentuk yang berbeda dari mioglobin non-teroksigenasi dan mutasi ini tidak mempengaruhi yang tetangga.
Wacana ini berbeda dalam hal protein terkait seperti hemoglobin: ketika sebuah rantai diberi oksigen, ia diinduksi untuk mengubah bentuknya tetapi modifikasi ini bersifat tiga dimensi sehingga rantai tetrameter lainnya juga terpengaruh. Fakta bahwa rantai-rantai tersebut saling terkait satu sama lain membuat orang berpikir bahwa modifikasi satu mempengaruhi yang lain bahkan jika pada tingkat yang berbeda; ketika sebuah rantai dioksigenasi, rantai tetrameter lain mengambil "sikap yang kurang bermusuhan" terhadap oksigen: kesulitan dengan mana rantai oksigenat berkurang ketika rantai di sebelahnya mengoksigenasi diri mereka sendiri. Argumen yang sama berlaku untuk deoksigenasi.
Struktur kuaterner dari deoxyhemoglobin disebut bentuk T (tense) sementara itu oxyhemoglobin disebut bentuk R (dirilis); dalam keadaan terentang ada serangkaian interaksi elektrostatik yang agak kuat antara asam amino asam dan asam amino basa yang mengarah pada struktur deoksihemoglobin yang kaku (inilah sebabnya "bentuk tegang"), sedangkan ketika oksigen terikat, entitas ini interaksi menurun (itulah sebabnya "formulir dirilis"). Lebih lanjut, dengan tidak adanya oksigen, muatan histidin (lihat struktur) distabilkan oleh muatan berlawanan dari asam aspartat sementara, di hadapan oksigen, ada kecenderungan protein kehilangan proton; semua ini berarti bahwa hemoglobin teroksigenasi adalah asam yang lebih kuat daripada hemoglobia terdeoksigenasi: efek bohr .
Tergantung pada pH, gugus heme lebih atau kurang mudah berikatan dengan oksigen: dalam lingkungan asam hemoglobin melepaskan oksigen lebih mudah (bentuk tegang stabil) sementara, dalam lingkungan dasar, ikatan dengan oksigen lebih kuat.

Setiap hemoglobin melepaskan 0, 7 proton per mol oksigen yang masuk (O2).
Efek Bohr memungkinkan hemoglobin meningkatkan kemampuannya untuk membawa oksigen.
Hemoglobin yang melakukan perjalanan dari paru-paru ke jaringan harus seimbang dengan tekanan, pH, dan suhu.
Kami melihat efek suhu .
Suhu di alveoli paru sekitar 1-1, 5 ° C lebih rendah dari suhu luar, sedangkan di otot suhu sekitar 36, 5-37 ° C; ketika suhu meningkat, faktor saturasi berkurang (dengan tekanan yang sama): ini terjadi karena energi kinetik meningkat dan disosiasi disukai.

Ada faktor lain yang dapat mempengaruhi kemampuan hemoglobin untuk mengikat oksigen, salah satunya adalah konsentrasi 2, 3 bisphosphoglycerate .
2, 3 bisphosphoglycerate adalah agen metabolik yang terdapat dalam eritrosit dalam konsentrasi 4-5 mM (tidak ada bagian tubuh lain yang hadir dalam konsentrasi tinggi).
Pada pH fisiologis, 2, 3 bisphosphoglycerate dideprotonasi dan memiliki lima muatan negatif; ia pergi ke irisan antara dua rantai beta hemoglobin karena rantai ini memiliki konsentrasi muatan positif yang tinggi. Interaksi elektrostatik antara rantai beta dan rantai bisphosphoglycerate memberikan kekakuan tertentu terhadap sistem: struktur tegang diperoleh yang memiliki sedikit afinitas terhadap oksigen; selama oksigenasi, maka, 2, 3 bisphosphoglycerate dikeluarkan.
Dalam eritrosit ada alat khusus yang mengubah 1, 3 bisphosphoglycerate (diproduksi oleh metabolisme) menjadi 2, 3 bisphosphoglycerate sehingga mencapai konsentrasi 4-5 mM dan oleh karena itu hemoglobin mampu menukar oksigen dalam jaringan.
Hemoglobin yang tiba di suatu jaringan berada dalam keadaan dilepaskan (terikat pada oksigen), tetapi di sekitar jaringan, ia mengalami karboksilasi dan masuk ke keadaan terentang: protein dalam keadaan ini memiliki kecenderungan lebih rendah untuk berikatan dengan oksigen, dibandingkan dengan keadaan yang dilepaskan, oleh karena itu hemoglobin melepaskan oksigen ke jaringan; lebih jauh lagi, karena reaksi antara air dan karbon dioksida, terdapat produksi ion H +, oleh karena itu oksigen lebih lanjut karena efek bohr.
Karbon dioksida berdifusi ke dalam eritrosit yang melewati membran plasma; mengingat bahwa eritrosit membentuk sekitar 40% dari darah, kita harus berharap bahwa hanya 40% karbon dioksida yang berdifusi dari jaringan yang masuk ke dalamnya, pada kenyataannya 90% karbon dioksida memasuki sel-sel merah karena mengandung enzim yang mengkonversi karbon dioksida dalam asam karbonat, maka konsentrasi karbon dioksida stasioner dalam eritrosit rendah dan karenanya kecepatan masuknya tinggi.

Fenomena lain yang terjadi ketika eritrosit mencapai jaringan adalah sebagai berikut: dengan gradien, HCO3- (turunan karbon dioksida) keluar dari eritrosit dan, untuk menyeimbangkan keluarnya muatan negatif, kami memiliki masuknya klorida yang menentukan peningkatan tekanan osmotik: untuk menyeimbangkan variasi ini ada juga masuknya air yang menyebabkan pembengkakan eritrosit (efek HAMBURGER). Fenomena sebaliknya terjadi ketika eritrosit mencapai alveoli paru: terjadi deflasi eritrosit (efek HALDANE). Jadi, eritrosit vena (langsung ke paru-paru) lebih bulat daripada yang arteri.